Research
The wound response in tomato – a paradigm for plant stress responses.
Life is dangerous ... especially for a plant that cannot simply run away to escape harmful environments. Plants are attacked by pests such as insects, nematodes, fungi, bacteria and viruses. Furthermore, they are exposed to abiotic forms of stress like drought, heat, cold, high salinity, and ultraviolet light. Unfavorable environmental conditions shift equilibria in ecosystems and result in enormous reductions in crop productivity. To cope with stress, plants have evolved remarkable defensive and protective strategies that allow them to develop into fertile organisms. Plant defense responses are continuously evolving in an evolutionary battle with pests, which in turn modify their invasive strategies to consume plants. This gave rise to an impressive arsenal of plant weapons. Many of the thousands of known plant secondary metabolites are thought to function in plant defense. To recognize danger and induce defenses, plants evolved a complex signaling network that responds to signals from the environment and regulates the synthesis of defense compounds.
The Systemic Wound Response
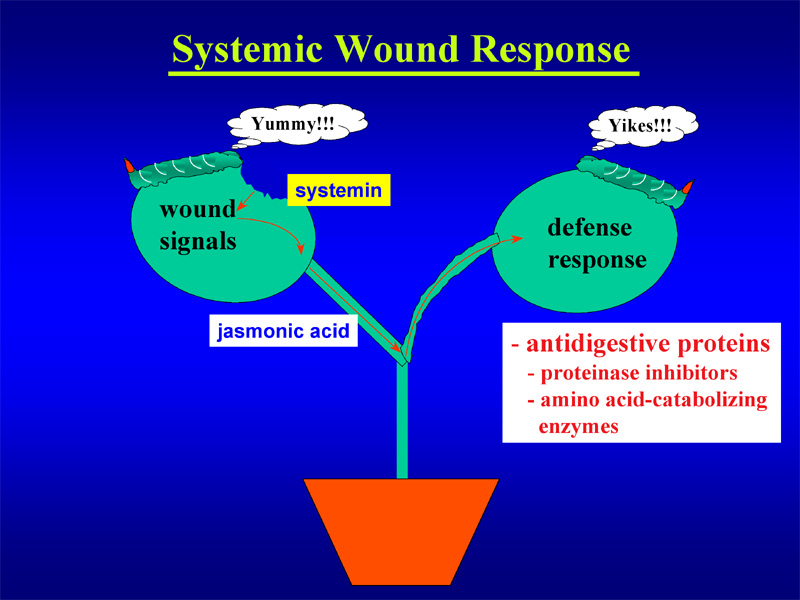
My group is employing tomato as a model plant. When tomato plants are attacked by chewing insects such as caterpillars, signals derived from these insects and the wounded plant are generated that can be perceived by plant cells. One of these wound signals is a small plant peptide called systemin which is thought to be processed from a precursor protein, called prosystemin. By binding to a membrane-bound receptor, systemin initiates an intracellular signal transduction pathway that relays and amplifies the wound signal leading to the activation of defense genes 1. Many plant defense genes encode antinutritive proteins such as proteinase inhibitors and enzymes that break down essential amino acids. When ingested, these proteins cause adverse effects on growth and development of the attacking insects by preventing uptake of essential amino acids in the insect gut 2. In addition, systemin activates indirect plant defenses through the release of volatile compounds 3,4. These volatiles are also released when plants are attacked by herbivorous insects and function as a ‘cry for help’. They serve as cues for predators or parasitoids which home in on the source of these volatiles to locate the herbivore and destroy it. An important signaling molecule synthesized in response to systemin and other wound signals is jasmonic acid. This plant hormone acts not only as a second messenger within the cells, but also as a long-distance signal that moves within the plant to distant leaves where it induces the wound response5.
Signal perception and signal transduction through MAPK pathways
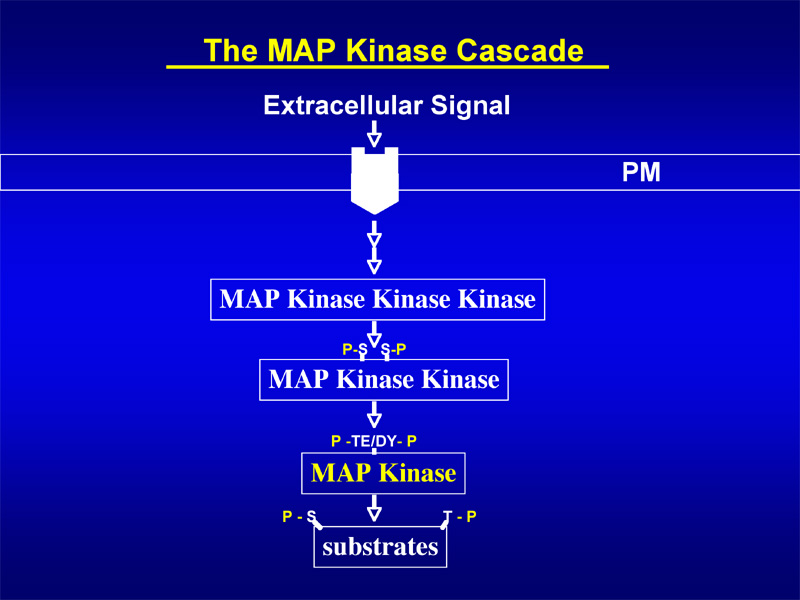
The first step for a successful stress response is the sensing or perception of stress signals by plant cells. The perceived signal is then relayed via a chain of events that leads to an amplification of the signal, and eventually to gene activations and a cellular response. Mitogen-activated protein kinases (MAPKs or MPKs) are important elements in eukaryotic stress signal transduction networks. A MAPK is part of the 'MAPK cascade' which consists of three functionally linked kinases. MAPKKKs can be activated by extracellular signals. In turn, they activate MAPKKs by phosphorylation, which then phosphorylate and activate MAPKs. MAPK cascades regulate the activity of transcription factors or cytosolic enzymes. The Arabidopsis genome contains ~ 60 putative MAPKKKs, only 10 MAPKKs, and 20 MAPKs.
Research projects
Our aim is to characterize the molecular mechanisms of stress signal transduction, and the highly complex plant signaling network that enables a plant cell to integrate multiple stress signals and launch an appropriate cellular defense response. The focus is on plant-insect interactions.
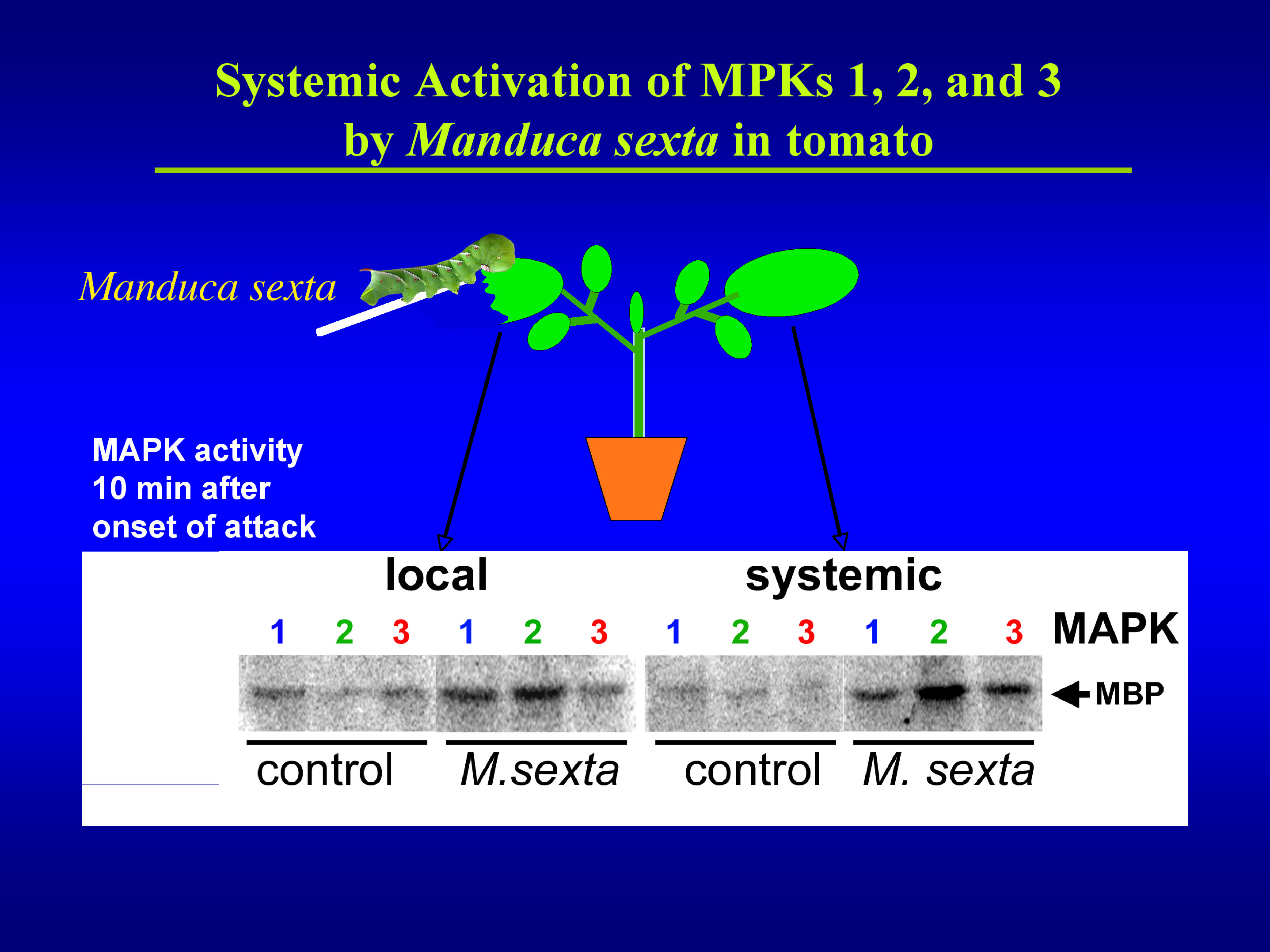
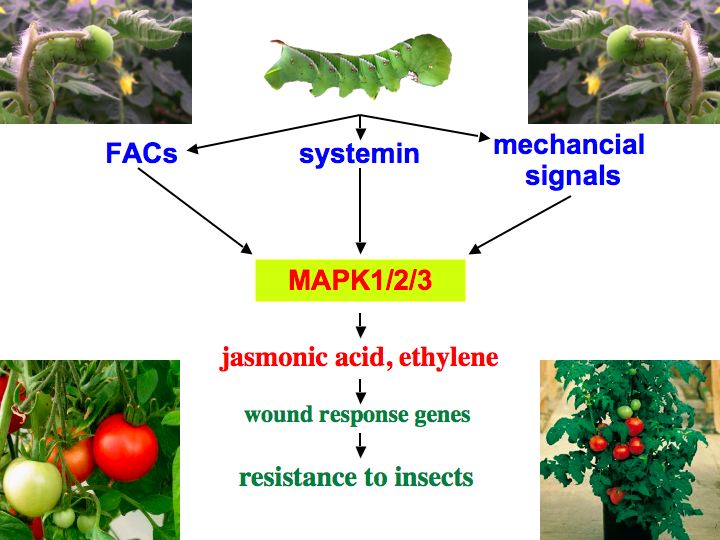
We identified three specific MAPKs that respond to herbivorous insects, mechanical wound signals, systemin, oligosaccharide elicitors and ultraviolet-B (UV-B) radiation 6,7,8. UV-B radiation does not fully induce the wound response. However, when cells were irradiated with UV-B and subsequently wounded, the accumulation of defense proteins was increased in a synergistic manner when compared to non-irradiated plants 9. This provides an excellent system to study how a plant cell integrates more than one stress signal to activate a cellular defense response. Moreover, it indicates that changes in the global environment such as increased ground level UV-B radiation due to stratospheric ozone depletion (ozone hole) might have unexpected consequences for plant-consumer interactions 10.
By employing virus-induced gene silencing (VIGS) to silence the three tomato MAPK genes MPK1, 2, and 3, we demonstrated that these MAPKs regulate jasmonic acid and ethylene synthesis in response to systemin and wounding. Since synthesis of these plant hormones is required for defense gene activation, the silenced plants showed not only a severely impaired wound response but also a strongly reduced resistance against herbivorous insects 8. Chewing insects generate other MAPK-activating signals in addition to systemin, such as mechanical signals 8,11 and fatty acid-amino acid conjugates (FACs) 12. The corresponding signaling pathways all converge on the same three MAPKs, which explains why silencing of these MAPKs prevented a successful defense response against the attacking insect larvae.

Projects in the Stratmann group are concerned with the systemin receptor, the MAPK cascade, and the COP9 signalosome.
One project is focused on the characterization of the systemin receptor SR160. A line of evidence suggested that the systemin receptor is identical to the brassinosteroid receptor BRI1 13,14. However, this concept was refuted when it was shown that bri1 null mutants in tomato have a wild type-like response to systemin 15. Our results are consistent with these findings but show that BRI1, when overexpressed in tobacco cells, can function as a systemin binding protein. However, binding of systemin to BRI1 does not relay the signal into the cell and does not trigger a defense response 16,17. It will now be important to identify the elusive systemin receptor.
Other projects revolve around MAPK pathways. Since MAPKs and MAPKKs function as convergence points for multiple stress signals, it is critical to understand how they contribute to signaling specificity in MAPK pathways. Although there are many MAPKKs and MAPKs, only a few of them seem to be involved in stress responses. Moreover, these few stress-responsive MAPKs respond to almost any stress signal. How can signaling components that are shared by multiple input signals lead to signal-specific responses? Regulation of the MAPK cascade at the level of all three kinases is likely to contribute to signaling specificity. We study the role of MAPKKs and MAPKs in plant responses to herbivory and other stresses, and how they tie into the stress signaling network.
In a new NSF-funded project, my group studies the COP9 signalosome, a multiprotein complex consisting of 8 subunits (CSN1-8). We have shown that the CSN5 protein operates as part of a molecular switch that determines whether defense responses to insects or pathogens will be activated. Partial knock-out of the CSN5 gene resulted in a strongly reduced resistance of tomato plants against herbivorous insects and upregulation of pathogenesis-related genes 18.
In the long run, our studies will provide clues for biotechnology on how to alter signaling networks in crop plants to make them more resistant to the stresses they encounter in an agricultural environment.
References:
- Ryan, C.A. (2000) The systemin signaling pathway: differential activation of plant defensive genes. Biochim. Biophys. Acta, 1477, 112-121.
- Chen, H., Wilkerson, C.G., Kuchar, J.A., Phinney, B.S. and Howe, G.A. (2005) Jasmonate-inducible plant enzymes degrade essential amino acids in the herbivore midgut. PNAS, 102, 19237-19242.
- Corrado, G., Sasso, R., Pasquariello, M., Iodice, L., Carretta, A., Cascone, P., Ariati, L., Digillio, M.C., Guerrieri, E., Rao, R., 2007. Systemin regulates both systemic and volatile signaling in tomato plants. J. Chem. Ecol. 33, 669-681.
- Degenhardt, D.C., Hind., S.R., Stratmann, J.W., and Lincoln, D.E. (2010) Systemin and jasmonic acid regulate constitutive and herbivore-induced systemic volatile emissions in tomato, Solanum lycopersicum. In revision (2010).
- Howe, G.A. (2004) Jasmonates as Signals in the Wound Response. J. Plant Growth Regul., 23, 223 - 237.
- Holley, S.R., Yalamanchili, R.D., Moura, S.D., Ryan, C.A. and Stratmann, J.W. (2003) Convergence of signaling pathways induced by systemin, oligosaccharide elicitors, and ultraviolet-B radiation at the level of mitogen-activated protein kinases in Lycopersicon peruvianum suspension-cultured cells. Plant Physiol., 132, 1728-1738.
- Higgins, R., Lockwood, T., Holley, S., Yalamanchili, R. and Stratmann, J. (2007) Changes in extracellular pH are neither required nor sufficient for activation of mitogen-activated protein kinases (MAPKs) in response to systemin and fusicoccin in tomato. Planta, 225, 1535-1546.
- Kandoth, P.K., Ranf, S., Pancholi, S.S., Jayanty, S., Walla, M.D., Miller, W., Howe, G.A., Lincoln, D.E. and Stratmann, J.W. (2007) Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects. PNAS, 104, 12205-12210.
- Stratmann, J.W., Stelmach, B.A., Weiler, E.W. and Ryan, C.A. (2000) UVB/UVA radiation activates a 48 kDa myelin basic protein kinase and potentiates wound signaling in tomato leaves. Photochem. Photobiol., 71, 116-123.
- Stratmann, J.W. (2003) Ultraviolet-B (UV-B) radiation co-opts defense signaling pathways. Trends Plant Sci., 11, 526-533.
- Stratmann, J.W. and Ryan, C.A. (1997) Myelin basic protein kinase activity in tomato leaves is induced systemically by wounding and increases in response to systemin and oligosaccharide elicitors. PNAS, 94, 11085-11089.
- Wu, J., Hettenhausen, C., Meldau, S. and Baldwin, I.T. (2007) Herbivory Rapidly Activates MAPK Signaling in Attacked and Unattacked Leaf Regions but Not between Leaves of Nicotiana attenuata. Plant Cell, 19, 1096-1122.
- Scheer, J. and Ryan, C.A. (2002) The systemin receptor SR160 from Lycopersicon peruvianum is a member of the LRR receptor kinase family. PNAS, 99, 9585-9590.
- Montoya, T., Nomura, T., Farrar, K., Kaneta, T., Yokota, T. and Bishop, G. J. (2002) Cloning the tomato curl3 gene highlights the putative dual role of the leucine-rich repeat receptor kinase tBRI1/SR160 in plant steroid hormone and peptide hormone signaling. Plant Cell, 14, 3163-3176.
- Holton, N., Cano-Delgado, A., Harrison, K., Montoya, T., Chory, J. and Bishop, G.J. (2007) Tomato BRASSINOSTEROID INSENSITIVE1 Is Required for Systemin-Induced Root Elongation in Solanum pimpinellifolium but Is Not Essential for Wound Signaling. Plant Cell, 19: 1709-1717.
- Malinowski, R., Higgins, R., Luo, Y., Piper, L., Nazir, A., Bajwa, V., Clouse, S.D., Thompson, P.R., Stratmann, J.W. (2009) The tomato brassinosteroid receptor BRI1 increases binding of systemin to tobacco plasma membranes, but is not involved in systemin signaling. Plant Mol. Biol. 70:603-616.
- Hind, S.R., Malinowski, R., Yalamanchili, R., and Stratmann. J.W. (2010) Tissue-type specific systemin perception and the elusive systemin receptor. Plant Signaling & Behavior 5,1: 41-44.
- Hind R.S., Pulliam, S.E., Veronese, P., Shantharaj, D., Nazir, A., Jacobs, N.S., Stratmann, J.W. (2010) The COP9 signalosome controls jasmonic acid synthesis and plant responses to herbivory and pathogens. In review.

